
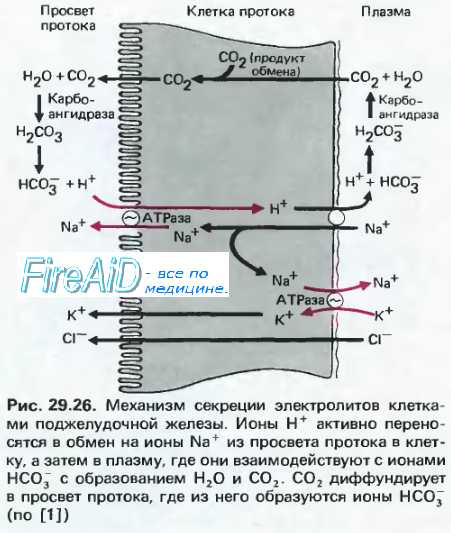
Нервно гуморальная регуляция поджелудочной железы

Оглавление темы “Сокращение желудка. Пищеварительные функции поджелудочной железы. Функции печени.”: Нервная регуляция секреторной функции поджелудочной железы. Гуморальная ( гормональная ) регуляция секреции поджелудочной железы.Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. В привычных условиях приема пищи панкреатическое сокоотделение начинается уже на вид, запах пищи и другие раздражители, сопутствующие приему пищи, что свидетельствует об условно-рефлекторном механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов. Выделяющийся при возбуждении этих окончаний ацетилхо-лин активирует М-холинорецепторы постсинаптической мембраны. При этом освобождаются вторичные посредники (Са++ и ГЦ-цГМФ), которые и вызывают секреторную деятельность панкреоцитов и эпителиальных клеток протоков. При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка, что приводит к продолжению безусловно-рефлекторного сокоотделения поджелудочной железой, вызванного стимуляцией рецепторов ротовой полости. Поступление в двенадцатиперстную кишку желудочного содержимого вызывает наряду с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастро-интестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой. Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин, выделяемый У-клетками слизистой оболочки антрального отдела желудка. Возбуждающее влияние этих гормонов подкрепляется бомбезином, серотонином и инсулином. Тормозят выделение сока соматостатин, глюкагон, энкефалины, вещество Р, ТИП, ПП, а также кальцитонн и АКТГ. Но их роль в комплексной гуморальной регуляции поджелудочной железы изучена недостаточно. – Также рекомендуем “Секреция сока поджелудочной железы. Фазы ( этапы ) секреции сока поджелудочной железы.” |
Источник
Регуляция секреции поджелудочной железы осуществляется комплексом нейрогуморальных механизмов.
Различают три фазы секреции: главное, желудочную и кишечную. Во время главной фазы секреции основная роль принадлежит нервным влияниям, которые реализуются через блуждающий нерв при условно и безумовнорефлекторних реакций. Под влиянием вида, запаха пищи, ее поступления в ротовую полость рефлекторно выделяется сок поджелудочной железы. Секреция начинается уже через 1-2 мин после начала приема пищи. В это время выделяется умеренное количество ферментов. Сок содержит незначительное количество воды и электролитов. Симпатические нервы осуществляют трофическое влияние на поджелудочную железу. их импульсы усиливают синтез органических веществ, в то же время подавляя их выделения. Поэтому эмоции и другие состояния, в результате которых возбуждается симпатический отдел вегетативной нервной системы, тормозят выделение сока.
При желудочной фазы нервные влияния сохраняются, но начинают действовать гуморальные факторы, в частности желудочный гастрин.
Кишечная фаза характеризуется четкой зависимостью количества сока и его состав от состава химуса. В настоящее время решающее значение имеют гуморальные факторы. Под влиянием химуса, поступившего в двенадцатиперстную кишку, образуются два гормона – секретин и ХЦК-ПЗ. То секретин образуется в S-клетках слизистой оболочки двенадцатиперстной кишки под влиянием НС1, ХЦК-ПЗ, в I-клетках этой оболочки – под влиянием продуктов гидролиза белков и жиров.
Секретин действует на клетки протоков поджелудочной железы. Под его влиянием выделяется много сока с высокой концентрацией гидрокарбонатов и малым количеством ферментов.
ХЦК-ПЗ влияет на синтез и выделение ферментов ацинарных клеток железы. В настоящее время выделяется мало сока, но он содержит значительное количество ферментов. На функцию ацинарных клеток влияют также гормоны собственно поджелудочной железы (есть данные, свидетельствующие о том, что кровь вначале протекает через панкреатические островки (островки Лангерганса) и лишь затем достигает ацинарных клеток.
Под действием секретина происходит щелочная реакция в тонкой кишке. Секретин начинает выделяться в кровь, когда рН в двенадцатиперстной кишке уменьшается до 4,5. При рН меньше от 3,0 выделения секретина значительно возрастает. Тогда выделяется сок с высокой концентрацией гидрокарбонатов. Он нейтрализует кислую реакцию химуса, поступающего из желудка. Таким образом, кислой будет реакция химуса, тем активнее она нейтрализуватиметься.
Если в химусе много белков или жиров, то образуется значительное количество ХЦК-ПЗ. Сок поджелудочной железы в этих условиях содержать высокоактивные ферменты, что обеспечит полноценный гидролиз названных веществ.
Влияние на ацинарных клетках реализуется через фосфолипазу С, а на клетки протоков – через цАМФ.
Основные стимуляторы секреции железы ацетилхолин, гастрин, секретин и ХЦК-ПЗ взаимодействуют между собой и усиливают конечный результат – они потенцируя влияние.
При приеме пищи с различным содержанием белков, жиров и углеводов
изменяются количество и состав сока. Таким образом поджелудочная железа приспосабливается к различным условиям, то есть происходит ее адаптация.
Источник
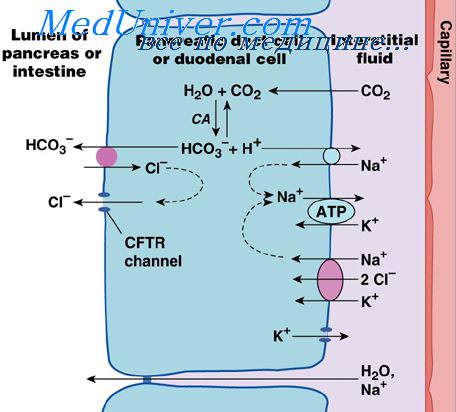
Регуляция секреции поджелудочной железы. Этапы панкреатической секрецииПоявление панкреатической секреции обусловлено тремя важными основными стимулами. Первые два стимула (ацетилхолин и холецистокинин) стимулируют ацинарные клетки поджелудочной железы, вызывая продукцию большого количества панкреатических пищеварительных ферментов и, соответственно, небольших количеств воды и электролитов, сопутствующих ферментам. Без воды большинство ферментов временно сохраняются в ацинусах и протоках до тех пор, пока не образуется более жидкий секрет, который может поступить в двенадцатиперстную кишку. Секретин в противоположность первым двум основным стимулам вызывает секрецию эпителием протоков поджелудочной железы большого количества водного раствора бикарбоната натрия. Усиливающие эффекты различных стимулов. Когда различные стимулы панкреатической секреции возникают одновременно, общая секреция гораздо больше, чем сумма секретов, вызванных каждым из них по отдельности, поэтому говорят, что разные стимулы усиливают друг друга. Таким образом, панкреатическая секреция является обычно результатом действия не одного, а комбинированных влияний перемноженных основных стимулов.
Фазы панкреатической секрецииПанкреатическая секреция аналогично желудочной секреции проходит три фазы: мозговую, желудочную и кишечную. В течение желудочной фазы продолжается нервная стимуляция ферментной секреции, составляя еще другие 5-10% панкреатических ферментов, выделяемых после приема пищи. Но опять только малые количества достигают двенадцатиперстной кишки из-за продолжающегося недостатка значимой секреции жидкости. Кишечная фаза. После того, как химус покидает желудок и поступает в тонкий кишечник, панкреатическая секреция становится обильной, главным образом в ответ на гормон секретин. Секретин стимулирует секрецию значительного количества ионов бикарбоната. Нейтрализация кислого химуса из желудка. Секретин представляет собой полипептид, содержащий 27 аминокислот (молекулярная масса около 3400). Он присутствует в слизистой двенадцатиперстной кишки и тощей кишки в неактивной форме просекретина в так называемых S-клетках. Когда кислый химус с рН менее 4,5-5,0 поступает в двенадцатиперстную кишку из желудка, это вызывает выделение дуоденальный слизи и активацию секретина, который затем абсорбируется в кровь. Единственной действительно важной составляющей химуса является соляная кислота желудка, которая вызывает выброс секретина. Секретин, в свою очередь, вызывает секрецию поджелудочной железой большого количества жидкости, содержащей высокую концентрацию иона бикарбоната (вплоть до 145 мэкв/л), но низкую концентрацию ионов хлора. Механизм образования секретина является особенно важным по двум причинам: (1) секретин начинает высвобождаться из слизистой тонкого кишечника, когда рН дуоденального содержимого падает ниже 4,5-5,0; (2) выброс секретина резко увеличивается, когда рН падает до 3,0. Это немедленно вызывает обильную секрецию панкреатического сока, содержащего существенное количество бикарбоната натрия. Конечным итогом является следующая реакция в двенадцатиперстной кишке: Затем угольная кислота немедленно диссоциирует на двуокись углерода и воду. Углекислый газ абсорбируется в кровь и выводится через легкие, оставляя нейтральный раствор хлорида натрия в двенадцатиперстной кишке. Таким способом немедленно нейтрализуется кислое содержимое, опорожняющееся в двенадцатиперстную кишку из желудка. Слизистая тонкой кишки не может противостоять переваривающему действию кислого желудочного сока, и это является существенным защитным механизмом, предотвращающим развитие дуоденальных язв. Секреция иона бикарбоната поджелудочной железой обеспечивает подходящий рН для работы панкреатических пищеварительных ферментов, действие которых оптимально в слабощелочной или нейтральной среде при рН от 7,0 до 8,0. К счастью, рН секреции натрия бикарбоната в среднем равняется 8,0. Холецистокинин и его вклад в регуляцию секреции пищеварительных ферментов поджелудочной железы. Присутствие пищи в верхнем отделе тонкого кишечника вызывает также выделение из I-клеток, находящихся в слизистой двенадцатиперстной кишки и верхней части тощей кишки, второго гормона — холецистокинина. Это полипептид, содержащий 33 аминокислоты. Такое выделение холецистокинина происходит в результате присутствия протеоз, пептонов (продуктов частичного переваривания белков) и длинноцепочечных жирных кислот в химусе, поступающем из желудка. Холецистокинин, как и секретин, с кровью доставляется к поджелудочной железе, но вместо стимуляции секреции бикарбоната натрия обусловливает преимущественное выделение ацинарными клетками пищеварительных ферментов, причем в больших количествах. Этот эффект подобен тому, который вызывает вагусная стимуляция, но только он более выраженный и составляет от 70 до 80% общей панкреатической секреции ферментов после приема пищи. Различия между эффектами панкреатической стимуляции секретином и холецистокинином показаны на рисунке, который демонстрирует: (1) интенсивную секрецию бикарбоната натрия в ответ на кислоту в двенадцатиперстной кишке, стимулированную секретином; (2) двоякий эффект в ответ на жир (мыла); (3) сильную пищеварительную секрецию ферментов, вызванную холецистокинином (когда пептоны поступают в двенадцатиперстную кишку). – Также рекомендуем “Физиология секреции желчи. Физиологическая анатомия секреции желчи” Оглавление темы “Пищеварительные соки. Переваривание углеводов, белков, жиров”: |
Источник
Слабое сокоотделение натощак резко
усиливается во время (через 2— 3 мин) и
после приема пищи. Панкреатическое
сокоотделение начинается уже на вид,
запах пищи как условно-рефлекторный
механизме секреции. При раздражении
пищевыми веществами рецепторов слизистой
оболочки ротовой полости, глотки и
пищевода возникает безусловно-рефлекторное
отделение панкреатического сока. Нервные
импульсы от раздражаемых рецепторов
по афферентным путям достигают бульбарного
центра панкреатической секреции, где
они переключаются на преганглионарные
нейроны ядер блуждающего нерва, по
эфферентным волокнам которого достигают
постганглионарных нейронов. Их аксоны
образуют синаптические окончания на
базальных мембранах панкреацитов.
Выделяющийся при возбуждении этих
окончаний ацетилхолин активирует
М-холинорецепторы постсинаптической
мембраны.
При раздражении чревных нервов,
осуществляющих симпатическую иннервацию
поджелудочной железы, ее сокоотделение
тормозится (за счет активации
р-адренорецепторов медиатором
норадреналином). Но торможение секреции
сопровождается накоплением секреторного
материала в панкреацитах.
Поступление пищи в желудок во время
приема пищи вызывает раздражение хемо-
и механорецепторов желудка.
Поступление в двенадцатиперстную кишку
желудочного содержимого вызывает вместе
с дуоденопанкреатическим секреторным
рефлексом (с рецепторов слизистой
оболочки) выделение из эндокринных
клеток гастро-интестинальных гормонов,
стимулирующих или тормозящих сокоотделение
поджелудочной железой.
Роль основных гуморальных регуляторов
выполняют секретин и ХЦК, которые
вырабатываются S- и ССК-клетками слизистой
оболочки двенадцатиперстной кишки.
Секретин и ХЦК усиливают влияние друг
друга на панкреоциты, особенно на фоне
выделения ацетилхолина в синаптических
окончаниях холинергических нервных
волокон, иннервирующих железу. Важным
гуморальным возбудителем панкреатической
секреции является гастрин.
4) Изменится ли амплитула потенциала действия нервной клетки, если на нее воздейсвовать стимулами сверхпороговой силы
это возбудимые ткани, амплитуда не
измен: так как действует закон «Все или
ничего»
Билет № 5
1.потенциал действия,с графиком,
возбудимость,мера возбудимости,
Потенциал
действия (ПД) – быстрое изменение
мембранного потенциала в ответ на
действия раздражителя пороговой силы.
ПД имеет стандартные амплитуду и
временные параметры, не зависящие от
силы стимула – правило “ВСЕ ИЛИ НИЧЕГО”.
Начальная деполяризация мембраны под
действием раздражителя:

Если сила раздражителя достаточна,
чтобы деполяризовать мембрану до КУД,
открываются быстрые потенциал-зависимые
натриевые каналы. Клетка возбуждена –
возник нервный импульс.

Восстановление мембранного потенциала
покоя – реполяризация мембраны.

Следующий этап – восстановление
мембранного потенциала покоя –
реполяризация, обусловлена активным
ионным транспортом. Наиболее важен
процесс активного транспорта – это
работа Na/K- насоса, который выкачивает ионы натрия
из клетки, одновременно закачивая ионы
калия внутрь клетки. Восстановление
мембранного потенциала происходит
благодаря току ионов калия из клетки –
калиевые каналы активируются и пропускают
ионы калия до достижения равновесного
калиевого потенциала. Это процесс важен
потому, что до тех пор, пока не восстановлен
МПП, клетка не способна воспринимать
новый импульс возбуждения.
ГИПЕРПОЛЯРИЗАЦИЯ – кратковременное
увеличение МП после его восстановления,
которое обусловлено повышением
проницаемости мембраны для ионов калия
и хлора. Гиперполяризация бывает только
после ПД.
Деполяризация мембраны до КУД – могут
открыться любые натриевые каналы,
иногда кальциевые, и быстрые, и медленные,
и потенциал-зависимые, и рецептор-управляемые.
Это зависит от вида раздражителя и типа
клетокБыстрое поступление натрия в клетку –
открываются быстрые, потенциал-зависимые
натриевые каналы, и деполяризация
достигает точки реверса потенциала –
происходит перезарядка мембраны, знак
заряда меняется на положительный.Восстановление градиента концентрации
по калию – работа насоса. Калиевые
каналы активированы, калий переходит
из клетки во внеклеточную среду –
реполяризация, начинается восстановление
МППСледовая деполяризация, или отрицательный
следовой потенциал – мембрана еще
деполяризована относительно МПП.Следовая гиперполяризация. Калиевые
каналы остаются открытыми и дополнительный
ток калия гиперполяризует мембрану.
После этого клетка возвращается к
исходному уровню МПП. Длительность ПД
составляет для разных клеток от 1 до
3-4 мс.
Обратите внимание на три величины
потенциала, важные и постоянные для
каждой клетки ее электрические
характеристики.
МПП – электроотрицательность мембраны
клетки в покое, обеспечивающая способность
к возбуждению – возбудимость.КУД – критический уровень, величина
мембранного потенциала, при достижении
которой открываются быстрые, потенциал
зависимые натриевые каналы и происходит
перезарядка мембраны за счет поступления
в клетку положительных ионов натрия.
Чем выше электроотрицательность
мембраны, тем труднее деполяризовать
ее до КУД, тем менее возбудима такая
клетка.Точка реверса потенциала (овершут) –
такая величина положительного мембранного
потенциала, при которой положительно
заряженные ионы уже не проникают в
клетку – кратковременный равновесный
натриевый потенциал.
При действии раздражителя
подпороговой силы возникает неполная
деполяризация – ЛОКАЛЬНЫЙ ОТВЕТ (ЛО).
Неполная, или частичная деполяризация
– это такое изменение заряда мембраны,
которое не достигает критического
уровня деполяризации (КУД).
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник