
Секреция пищеварительного сока поджелудочной железы регулируется нервными и гуморальными путями

Оглавление темы “Сокращение желудка. Пищеварительные функции поджелудочной железы. Функции печени.”: Нервная регуляция секреторной функции поджелудочной железы. Гуморальная ( гормональная ) регуляция секреции поджелудочной железы.Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. В привычных условиях приема пищи панкреатическое сокоотделение начинается уже на вид, запах пищи и другие раздражители, сопутствующие приему пищи, что свидетельствует об условно-рефлекторном механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов. Выделяющийся при возбуждении этих окончаний ацетилхо-лин активирует М-холинорецепторы постсинаптической мембраны. При этом освобождаются вторичные посредники (Са++ и ГЦ-цГМФ), которые и вызывают секреторную деятельность панкреоцитов и эпителиальных клеток протоков. При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка, что приводит к продолжению безусловно-рефлекторного сокоотделения поджелудочной железой, вызванного стимуляцией рецепторов ротовой полости. Поступление в двенадцатиперстную кишку желудочного содержимого вызывает наряду с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастро-интестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой. Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин, выделяемый У-клетками слизистой оболочки антрального отдела желудка. Возбуждающее влияние этих гормонов подкрепляется бомбезином, серотонином и инсулином. Тормозят выделение сока соматостатин, глюкагон, энкефалины, вещество Р, ТИП, ПП, а также кальцитонн и АКТГ. Но их роль в комплексной гуморальной регуляции поджелудочной железы изучена недостаточно. – Также рекомендуем “Секреция сока поджелудочной железы. Фазы ( этапы ) секреции сока поджелудочной железы.” |
Источник
Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. Панкреатическое сокоотделение начинается уже на вид, запах пищи как условно-рефлекторный механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов.
Выделяющийся при возбуждении этих окончаний ацетилхолин активирует М-холинорецепторы постсинаптической мембраны.
При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка.
Поступление в двенадцатиперстную кишку желудочного содержимого вызывает вместе с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастро-интестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой.
Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин.
Изменится ли амплитула потенциала действия нервной клетки, если на нее воздейсвовать стимулами сверхпороговой силы
это возбудимые ткани, амплитуда не измен: так как действует закон «Все или ничего»
Билет № 5
1.потенциал действия,с графиком, возбудимость,мера возбудимости,
Потенциал действия (ПД) – быстрое изменение мембранного потенциала в ответ на действия раздражителя пороговой силы. ПД имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула – правило “ВСЕ ИЛИ НИЧЕГО”.
Начальная деполяризация мембраны под действием раздражителя:
Если сила раздражителя достаточна, чтобы деполяризовать мембрану до КУД, открываются быстрые потенциал-зависимые натриевые каналы. Клетка возбуждена – возник нервный импульс.
Восстановление мембранного потенциала покоя – реполяризация мембраны.
Следующий этап – восстановление мембранного потенциала покоя – реполяризация, обусловлена активным ионным транспортом. Наиболее важен процесс активного транспорта – это работа Na/K – насоса, который выкачивает ионы натрия из клетки, одновременно закачивая ионы калия внутрь клетки. Восстановление мембранного потенциала происходит благодаря току ионов калия из клетки – калиевые каналы активируются и пропускают ионы калия до достижения равновесного калиевого потенциала. Это процесс важен потому, что до тех пор, пока не восстановлен МПП, клетка не способна воспринимать новый импульс возбуждения.
ГИПЕРПОЛЯРИЗАЦИЯ – кратковременное увеличение МП после его восстановления, которое обусловлено повышением проницаемости мембраны для ионов калия и хлора. Гиперполяризация бывает только после ПД.
- Деполяризация мембраны до КУД – могут открыться любые натриевые каналы, иногда кальциевые, и быстрые, и медленные, и потенциал-зависимые, и рецептор-управляемые. Это зависит от вида раздражителя и типа клеток
- Быстрое поступление натрия в клетку – открываются быстрые, потенциал-зависимые натриевые каналы, и деполяризация достигает точки реверса потенциала – происходит перезарядка мембраны, знак заряда меняется на положительный.
- Восстановление градиента концентрации по калию – работа насоса. Калиевые каналы активированы, калий переходит из клетки во внеклеточную среду – реполяризация, начинается восстановление МПП
- Следовая деполяризация, или отрицательный следовой потенциал – мембрана еще деполяризована относительно МПП.
- Следовая гиперполяризация. Калиевые каналы остаются открытыми и дополнительный ток калия гиперполяризует мембрану. После этого клетка возвращается к исходному уровню МПП. Длительность ПД составляет для разных клеток от 1 до 3-4 мс.
Обратите внимание на три величины потенциала, важные и постоянные для каждой клетки ее электрические характеристики.
- МПП – электроотрицательность мембраны клетки в покое, обеспечивающая способность к возбуждению – возбудимость.
- КУД – критический уровень, величина мембранного потенциала, при достижении которой открываются быстрые, потенциал зависимые натриевые каналы и происходит перезарядка мембраны за счет поступления в клетку положительных ионов натрия. Чем выше электроотрицательность мембраны, тем труднее деполяризовать ее до КУД, тем менее возбудима такая клетка.
- Точка реверса потенциала (овершут) – такая величина положительного мембранного потенциала, при которой положительно заряженные ионы уже не проникают в клетку – кратковременный равновесный натриевый потенциал.
При действии раздражителя подпороговой силы возникает неполная деполяризация – ЛОКАЛЬНЫЙ ОТВЕТ (ЛО). Неполная, или частичная деполяризация – это такое изменение заряда мембраны, которое не достигает критического уровня деполяризации (КУД).
Дата добавления: 2016-11-18; просмотров: 991 | Нарушение авторских прав | Изречения для студентов
Читайте также:
Рекомендуемый контект:
Поиск на сайте:
© 2015-2020 lektsii.org – Контакты – Последнее добавление
Источник
Регуляция функции желез пищеварительного тракта. Нервная регуляция секреции желез
Парасимпатическая стимуляция. Раздражение окончаний парасимпатических нервов пищеварительного тракта, как правило, увеличивает уровень секреции пищеварительных желез. В первую очередь это касается желез верхней части тракта (иннервируемых языкоглоточным и блуждающим нервами), таких как слюнные железы, железы пищевода, железы желудка, а также поджелудочная железа и бруннеровы железы двенадцатиперстной кишки. Это также справедливо для некоторых желез дистального отдела толстой кишки, иннервируемых тазовыми парасимпатическими нервами. Секрецию в остальных отделах тонкой кишки и в первых двух третях толстой кишки осуществляют главным образом местные нервные и гормональные стимулы в каждом отделе пищеварительной трубки.
Симпатическая стимуляция. Раздражение симпатических нервов, идущих к желудочно-кишечному тракту, приводит к умеренному локальному увеличению секреции некоторых желез. Но симпатическая стимуляция также вызывает сужение сосудов, которые кровоснабжают железы. В результате симпатическая стимуляция может производить двойной эффект: (1) отдельная симпатическая стимуляция обычно незначительно увеличивает секрецию; (2) если парасимпатическая или гормональная стимуляция уже вызвала обильную секрецию, то наслаивающаяся симпатическая стимуляция обычно уменьшает секрецию, иногда значительно, главным образом из-за уменьшения кровоснабжения вследствие сужения сосудов.
Гормональная регуляция секреции желез. В желудке и кишечнике несколько различных гастроинтестинальных гормонов помогают регулировать объем и характер секреции. Эти гормоны высвобождаются из слизистой желудочно-кишечного тракта в ответ на присутствие пищи в полости пищеварительного тракта. Затем гормоны всасываются в кровь и достигают желез, где вызывают секрецию. Этот тип стимуляции особенно важен для увеличения выброса желудочного и панкреатического соков, когда пища поступает в желудок и двенадцатиперстную кишку. По своему химическому составу гастроинтестинальные гормоны представляют собой полипептиды либо их производные.

Основные механизмы, благодаря которым железистые клетки функционируют, неизвестны, но экспериментальные данные указывают на следующие принципы секреции.
1. Питательные вещества, необходимые для появления секреции, должны вначале диффундировать или активно транспортироваться кровью в капилляры основания железистой клетки.
2. Большое количество митохондрий, находящихся внутри железистой клетки возле ее основания, используют энергию окисления для получения АТФ.
3. Затем энергия АТФ наряду с необходимыми компонентами питательных веществ используется для синтеза органических секреторных веществ. Этот синтез осуществляется практически полностью в эндоплазматическом ре-тикулуме и комплексе Голъджи железистой клетки. Рибосомы, прикрепленные к ретику-луму, ответственны за построение секретиру-емых белков.
4. Секреторные материалы транспортируются через канальцы эндоплазматического рети-кулума, проходя приблизительно за 20 мин весь путь к пузырькам комплекса Гольджи.
5. В комплексе Гольджи материалы модифицируются, суммируются, концентрируются и высвобождаются в цитоплазму в форме секреторных пузырьков, которые запасаются на апикальном конце секреторной клетки.
6. Эти пузырьки сохраняются до тех пор, пока нервные или гуморальные сигналы не приведут к выталкиванию содержимого пузырьков через поверхность клетки. Вероятно, это происходит следующим образом: управляющий сигнал вначале увеличивает проницаемость клеточной мембраны для ионов кальция, и кальций поступает в клетку. Кальций, в свою очередь, обусловливает встраивание большого количества пузырьков в апикальную поверхность клеточной мембраны. Затем апикальная клеточная мембрана лопается, высвобождая содержимое пузырьков наружу. Этот процесс называют экзоцитозом.
– Также рекомендуем “Секреция воды и электролитов в ЖКТ. Физиология слизи желудочно-кишечного тракта”
Оглавление темы “Желудочная и поджелудочная секреция”:
1. Регуляция функции желез пищеварительного тракта. Нервная регуляция секреции желез
2. Секреция воды и электролитов в ЖКТ. Физиология слизи желудочно-кишечного тракта
3. Физиология слюны. Секреция слюны
4. Гигиенические свойства слюны. Нервная регуляция слюноотделения
5. Секреция в пищеводе. Физиология желудочной секреции
6. Физиология секреции пепсиногена. Секреция желудочной слизи и гастрина
7. Стимуляция секреции кислоты в желудке. Стимуляция секреции гастрина
8. Физиология и фазы желудочной секреции. Торможение и регуляция желудочной секреции
9. Строение гастрина. Физиология секреции поджелудочной железы
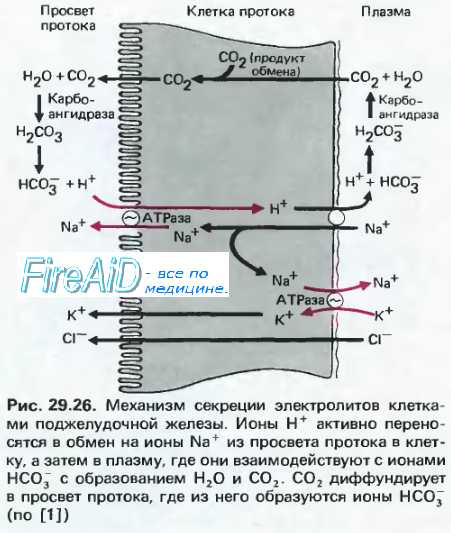
10. Предотвращение самопереваривания поджелудочной железы. Секреция ионов бикарбоната
Источник
Оглавление темы “Функция всасывания кишечника. Пищеварение в ротовой полости и функция глотания.”: Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.Центральные, периферические и местные рефлексы осуществляются в тесном взаимодействии с гуморальным механизмом регуляции миоцитов, гландулоцитов и нервных клеток. В слизистой оболочке желудочно-кишечного тракта и в поджелудочной железе имеются эндокринные клетки, которые вырабатывают гастроинте-стинальные гормоны (регуляторные пептиды, энтерины). Эти гормоны через кровоток и местно (паракринно, диффундируя через межклеточную жидкость) оказывают влияние на миоциты, гландулоциты, интрамураль-ные нейроны и эндокринные клетки. Их выработка запускается рефлекторно (через блуждающий нерв) во время приема пищи и длительное время поддерживается за счет раздражающего влияния продуктов гидролиза пищевых веществ и экстрактивных веществ. Таблица 11.1. Гормоны желудочно-кишечного тракта, место их образования и вызываемые ими эффекты
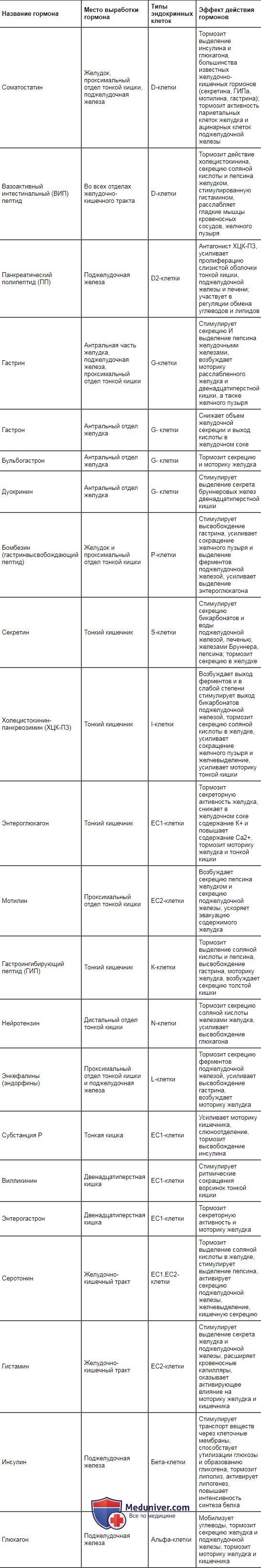 Место выработки основных гастроинтестинальных гормонов, вызываемые ими эффекты и продуцирующие их клетки представлены в табл. 11.1. В настоящее время обнаружено около 30 регуляторных пептидов. Как следует из представленной таблицы, они оказывают стимулирующее, тормозное и модулирующее влияние на секрецию пищеварительных соков, моторику гладкой мускулатуры желудочно-кишечного тракта, всасывание, выделение энтеринов эндокринными элементами слизистой оболочки желудка, кишечника и поджелудочной железы. Выделение гастроинтестинальных гормонов имеет каскадный характер. Например, под влиянием гастрина обкладочные клетки желез желудка увеличивают выработку соляной кислоты, которая в слизистой оболочке тонкой кишки стимулирует выделение S- и J-клетками секретина и холеци-стокинна — панкреозимина. Секретин усиливает секрецию воды и бикарбонатов поджелудочной железой и печенью, а холецистокинин — панкреозимин — возбуждает выделение ферментов поджелудочной железой и тормозит секрецию соляной кислоты обкладочными клетками, усиливает моторику тонкой кишки и желчного пузыря. Регуляторные пептиды, поступая в кровоток, быстро разрушаются в печени и почках и тем самым создают условия для осуществления эффектов других гастроинтестинальных гормонов. Выработка некоторых энтеринов носит циклический характер и может осуществляться и при отсутствии пищевого раздражителя. Например, мо-тилин, вырабатываемый ЕС2-клетками проксимального отдела тонкой кишки, вызывает сокращения мышц желудка и кишечника, совпадающие с периодами «голодной» активности пищеварительного тракта. – Также рекомендуем “Схема механизмов регуляции функций желудочно-кишечного тракта (жкт). Обобщенная схема механизмов регуляции функций пищеварительного тракта.” |
Источник