
Мейоза клетки поджелудочной железы

Эндокринная часть поджелудочной железы. Регенерация поджелудочной железы.
Островки состоят из эпителиальных клеток — панкреатических эндокриноцитов, или инсулоцитов. Величина островков, их форма и число входящих в состав клеток очень различны. Общее количество островков в поджелудочной железе достигает 1-2 млн. Средний размер островка 0,1-0,3 мм. Общий объем эндокринной части составляет около 3% всего объема железы. Островки пронизаны кровеносными капиллярами, окруженными перикапиллярным пространством. Эндотелий капилляров имеет фенестры, облегчающие поступление гормонов от инсулоцитов в кровь через перикапиллярное пространство.
В островковом эпителии различают 5 видов клеток: А-клетки, В-клетки, D-клетки, ВИП-клетки, РР-клетки.
А-клетки (альфа-клетки, или ацидофильные инсулоциты) — это крупные округлые клетки с бледным крупным ядром и цитоплазмой, содержащей ацидофильные гранулы. Гранулы обладают и аргирофилией. В состав этих гранул входит гормон глюкагон, расщепляющий гликоген и повышающий содержание сахара в крови.
А-клетки рассеяны по всему островку, образуя нередко небольшие скопления в центральной части. Они составляют около 20-25% от всех инсулоцитов.
В-клетки (бета-клетки, или базофильные инсулоциты) имеют кубическую или призматическую форму, крупное темное, богатое гетерохроматином ядро. Доля В-клеток достигает 70-75% от общего числа инсулоцитов. В цитоплазме В-клеток накапливаются осмиофильные гранулы, содержащие гормон инсулин. Инсулин регулирует синтез гликогена из глюкозы. При недостатке продукции инсулина глюкоза не превращается в гликоген, содержание ее в крови повышается и создаются условия для развития заболевания, называемого сахарным диабетом.
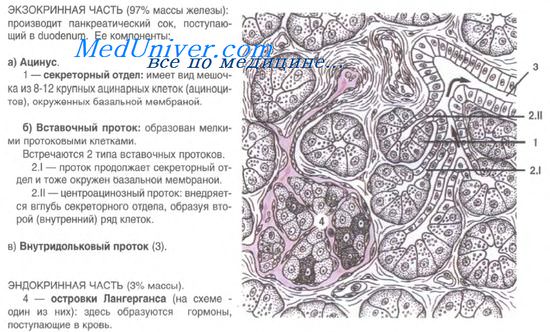
D-клетки (дельта-клетки, или дендритические инсулоциты) составляют 5-10% среди всех островковых клеток. Форма их иногда звездчатая с отростками. В цитоплазме определяются гранулы средних размеров и плотности. В гранулах накапливается гормон соматостатин. Он тормозит секрецию инсулина и глюкагона, снижает продукцию ряда гормонов желудочно-кишечного тракта — гастрина, секретина, энтероглюкагона, холецистокинина и др., подавляет секрецию соматотропного гормона в гипофизе.
ВИП-клетки (аргирофильные клетки) встречаются в островках в небольшом количестве. В цитоплазме выявляются плотные аргирофильные гранулы, содержащие вазоактиеный интестиналъный полипептид. Он обладает выраженным сосудорасширяющим дейтвием, снижает кровяное давление, угнетает секрецию соляной кислоты в желудке, стимулирует выделение глюкагона и инсулина.
РР-клетки — полигональной формы инсулоциты, расположенные преимущественно по периферии островка. Количество их — 2-5% от общего числа клеток островка. В цитоплазме РР-клеток выявляются мелкие гранулы, содержащие панкреатический полипептид. Основная роль панкреатического полипептида в организме — регуляция скорости и количества экзокринной секреции поджелудочной железы и желчи в печени. Таков клеточный состав островкового эпителия, представляющего собой мозаику дивергентно развивающихся клеточных дифферонов.
В дольках поджелудочной железы встречаются еще ацинозно-инсулярные клетки, в цитоплазме которых одновременно содержатся гранулы, характерные как для ацинозных, так и для островковых клеток.
Ткани поджелудочной железы иннервируют блуждающий и симпатический нервы. В интрамуральных вегетативных ганглиях находятся холинергические и пептидергические нейроны, волокна которых заканчиваются на клетках ацинусов и островков. Между нервными клетками ганглиев и островковыми клетками устанавливается тесная связь с образованием нейроинсулярных комплексов.
С возрастом в поджелудочной железе постепенно уменьшается количество островков. В островках наблюдаются закономерные возрастные изменения клеточных взаимоотношений, заключающиеся в быстрой смене после рождения преобладания А-клеток над В-клетками на преобладание В-клеток над А-клетками у взрослых. Затем происходит постепенное увеличение количества А-клеток, которое наряду с одновременным, хотя и незначительным, уменьшением числа В-клеток приводит нередко в пожилом и особенно старческом возрасте вновь к преобладанию А-клеток над В-клетками.
Регенерация поджелудочной железы. В эмбриогенезе островки растут благодаря пролиферации исходных клеток-предшественниц и их дивергентной дифференцировке в соответствующие клеточные диффероны. У взрослых физиологическая регенерация ацинозных и островковых клеток происходит в основном путем внутриклеточного обновления органелл. Митотическая активность клеток в связи с высокой специализацией низкая. После резекции части или повреждения органа наблюдается некоторое повышение уровня пролиферативной активности клеток ацинусов, протоков и островков, последующее образование новых ацинусов. Однако ведущей формой регенерации экзокринной части железы является регенерационная гипертрофия.
Восстановительные процессы в эндокринной части железы происходят за счет пролиферативной активности инсулоцитов и клеток эпителия протоков путем ацино-инсулярной трансформации.
– Также рекомендуем “Печень. Развитие печени. Строение печени.”
Оглавление темы “Строение желудка. Строение кишечника.”:
1. Пищевод. Слизистая пищевода. Строение стенки пищевода.
2. Желудок. Развитие желудка. Строение желудка. Железы желудка.
3. Эпителий желез желудка. Клетки желудка. Гормоны желудка.
4. Тонкая кишка. Развитие тонкой кишки. Строение тонкой кишки.
5. Эпителий тонкой кишки. Клетки тонкой кишки.
6. Толстая кишка. Развитие и строение толстой кишки. Червеобразный отросток.
7. Прямая кишка. Строение прямой кишки. Поджелудочная железа.
8. Эндокринная часть поджелудочной железы. Регенерация поджелудочной железы.
9. Печень. Развитие печени. Строение печени.
10. Гепатоциты. Строение гепатоцитов. Образование желчи.
Источник
Формирование поджелудочной железы у плода – эмбриогенез, морфогенез
Морфогенез поджелудочной железы человека начинается на 30-й день гестации. Формирование поджелудочной железы сначала сопровождается появлением на противоположных поверхностях головной кишки дорсальной и вентральной панкреатических почек. По мере увеличения эпителиальных почек происходит формирование древовидной системы протоков за счет их прогрессивного роста и ветвления.
Вследствие дальнейшего роста и вращения развивающейся кишечной трубки две панкреатические почки двигаются навстречу друг другу и сливаются на сроке гестации около 7 нед.
Сначала в железе определяются отдельные эндокринные клетки, а затем формируются островки поджелудочной железы. На сроке гестации около 20 нед в экзокринной части поджелудочной железы появляются признаки ферментативной активности. Собственно секреция начинается на 5-м месяце внутриутробного развития, причем продукция каждого из ферментов имеет индивидуальные особенности.
Развитие поджелудочной железы представляет собой один из классических примеров взаимодействия эпителия и мезенхимы. Было продемонстрировано, что для роста и дифференцировки поджелудочной железы необходимо присутствие мезенхимы, несмотря на то что как эндокринные, так и экзокринные клетки формируются из энтодермы головной кишки.
При изучении развития изолированных энтодермы и мезенхимы в различных условиях культивирования in vitro обнаружено, что «по умолчанию» идет дифференцировка поджелудочной железы, приводящая к образованию эндокринных клеток. В то же время для полноценного органогенеза железы необходимо совместное действие внеклеточного матрикса и мезенхимных факторов.
Дорсальная, или спинная, панкреатическая почка развивается в зоне, где экспрессия Shh подавляется факторами, продуцируемыми дорсальной областью хорды. Способность реагировать на регулирующие сигналы определяется изначальной экспрессией ряда энтодермальных факторов, таких как HNF1b, HNF3a иb (FoxAl и FoxA2), HNF4a, HNF6, GATA4, GATA5 и GATA6. Некоторые из указанных факторов контролируют экспрессию панкреатодуоденального гомеобокса 1 (Pdx-1).
Препанкреатическая энтодерма экспрессирует два уникальных гена — pdx-1 и blх-9. Одним из наиболее ранних признаков возникновения поджелудочной железы является экспрессия клетками панкреатической почки гена pdx-1. Клетки-предшественники, экспрессирующие ген pdx-1, дают начало всем клеткам поджелудочной железы.
Установлено, что различные факторы контролируют спецификацию дорсального и вентрального отделов поджелудочной железы, при этом экспрессия blx-9 в дорсальной препанкреатической энтодерме предшествует экспрессии pdx-1. У мышей при отсутствии Н1х-9 дорсальная препанкреатическая энтодерма не способна экспрессировать Pdx-1, вследствие чего не формируется дорсальная (спинная) панкреатическая почка.
Вентральная (брюшная) часть поджелудочной железы при этом характеризуется формированием всех четырех типов гормонпродуцирующих клеток островков Лангерганса, однако в существенно меньшем количестве.
Установлено, что белок PDX-1 вырабатывается в эпителии двенадцатиперстной кишки, расположенном в непосредственной близости к панкреатическим почкам, а также в эпителии самих панкреатических почек. У мышей с делецией гена pdx-1 при нормальном развитии всех отделов ЖКТ и всего тела в целом поджелудочная железа не формировалась.
Другая группа исследователей, независимо получивших мышей с нокаутом гена pdx-1, установила, что дорсальная почка поджелудочной железы формировалась, однако ее дальнейшее развитие прекращалось. Дефект развития, связанный с нокаутом гена pdx, ограничен эпителием, в то время как мезенхимные клетки в отсутствие pdx дифференцируются нормально.
Кроме того, у мышей с нокаутом гена pdx наиболее проксимальный отдел двенадцатиперстной кишки развивался аномально: в нем формировались везикулярные структуры, покрытые кубическим эпителием, а не ворсины с цилиндрическим эпителием. Это свидетельствует о том, что pdx-1 оказывает влияние на дифференцировку клеток на значительно большей площади, чем область, из которой развивается поджелудочная железа, что согласуется с ранее обозначенной зоной экспрессии указанного гена. Описана врожденная агенезия поджелудочной железы у человека, связанная с делецией единичного нуклеотида в гене pdx-1.
Сигнальный путь Notch определяет клетки, которые должны дифференцироваться в эндокринные. Предшественники эндокринных клеток временно экспрессируют нейрогенин (ngn) 3, который блокируется влиянием сигнального пути Notch. У мышей при отсутствии ngn3 исчезает способность к формированию любых эндокринных клеток.
С другой стороны, чрезмерная экспрессия ngn3 приводит к дифференцировке всей ткани поджелудочной железы в эндокринные клетки. Клетки, в которых экспрессия ngn3 блокируется сигнальным путем Notch, трансформируются в экзокринные. Для окончательной дифференцировки в экзокринные клетки необходимо присутствие дополнительного фактора — Р48.
У мышей с выключенной экспрессией Р48 отсутствует экзокринная часть поджелудочной железы, однако эндокринные островки полностью сформированы. Экспрессия ngn3 определяет экзокринную функцию клеток, однако период развития и характер взаимодействия с другими главными факторами, определяющими дифференцировку четырех основных типов эндокринных клеток, остаются неизученными. Более детальное описание современных представлений о молекулярных механизмах развития поджелудочной железы изложено в нескольких обзорах.
Учебное видео по развитию желудочно-кишечного тракта (эмбриогенезу)

– Также рекомендуем “Формирование тонкой кишки у плода – эмбриогенез, морфогенез”
Оглавление темы “Эмбриогенез желудочно-кишечного тракта”:
- Формирование желудочно-кишечного тракта у плода – эмбриология, морфогенез
- Регуляция специализации энтодермы при формировании желудочно-кишечного тракта у плода
- Генетическая регуляция формирования кишечной трубки
- Генетическая регуляция формирования органов желудочно-кишечного тракта
- Формирование пищевода у плода – эмбриогенез, морфогенез
- Формирование желудка у плода – эмбриогенез, морфогенез
- Формирование печени у плода – эмбриогенез, морфогенез
- Формирование поджелудочной железы у плода – эмбриогенез, морфогенез
- Формирование тонкой кишки у плода – эмбриогенез, морфогенез
- Дифференциация стволовых клеток тонкой кишки – эмбриогенез, морфогенез
Источник
Генетическая регуляция формирования поджелудочной железы человекаНесмотря на то что морфогенез поджелудочной железы был описан более 100 лет назад, молекулярные медиаторы, лежащие в основе этого процесса, были идентифицированы недавно. Современное понимание молекулярных механизмов развития поджелудочной железы предполагает совместное действие «обучающих» и «разрешающих» факторов в процессе формирования какого-либо органа. Если придерживаться этой точки зрения, то органогенез представляется удивительно постоянным явлением, воспроизводимым от животного к животному. Значит, вполне возможно действие многочисленных защитных и/или стимулирующих механизмов, суть которых пока не совсем понятна. Паренхиматозные клетки поджелудочной железы (как эндокринной, так и экзокринной частей) происходят из энтодермального эпителия примитивной кишечной трубки. Например, перед тем как зачаток поджелудочной железы будет эвагинировать (выпячиваться) из примитивной кишечной трубки, многие молекулярные программы, определяющие дальнейшее развитие поджелудочной железы, запускаются под действием соседних тканей. В процессе гаструляции энтодерма подразделяется на передний и задний домены под действием сигналов, исходящих из расположенной рядом мезэктодермы. Ретиноевая кислота, BMP и FGF играют важную роль в процессе поздней гаструляции, локализуя препанкреатический домен, который в дальнейшем будет отвечать за «обучающие» сигналы, определяющие развитие ткани поджелудочной железы. Будущие вентральная и дорсальная части поджелудочной железы подвергаются действию различных местных факторов, поэтому их индукция зависит от определенных моментов. Энтодерма, которая впоследствии будет дифференцироваться в вентральную часть поджелудочной железы, первой начинает экспрессировать панкреатические гены.
Этот процесс индуцируется активином и молекулами семейства BMP, которые происходят из расположенной рядом латеральной пластинки мезодермы. Формирование дорсальной части поджелудочной железы индуцируется другим механизмом. Ее энтодерма кратковременно контактирует сначала с нотохордой, а затем с дорсальной аортой. Контакт с нотохордой индуцирует локальное исключение Shh из препанкреатической энтодермы. Это сигнальное событие необходимо для развития дорсальной части поджелудочной железы, поскольку эктопическая экспрессия Shh влияет на последующие процессы. Контакт между энтодермой и нотохордой приводит к интерпозиции дорсальной аорты, что также обеспечивает возникновение значимых сигналов для последующего развития поджелудочной железы. Одним из них является фактор роста эндотелия сосудов. События, описанные ранее, вносят свой вклад в процесс формирования препанкреатической энтодермы, что важно для последующего образования ткани поджелудочной железы. Следующий шаг заключается в экспрессии факторов транскрипции в энтодерме и окружающей мезенхиме для дальнейшей дифференцировки этих клеток с целью последующего развития поджелудочной железы. Одним из значимых генов является blx-9, кодирующий протеин гомеобокса НВ9. Его генетическая аблация (отсутствие, обусловленное генетическими нарушениями) ведет к селективному отсутствию дорсальной части поджелудочной железы. Ген pdx-1, один из первых открытых факторов панкреатоспецифической транскрипции, экспрессируется как дорсальным, так и вентральным зачатками поджелудочной железы. Его аблация приводит к остановке развития железы сразу же после формирования этих структур. Было выявлено, что мутация гена pdx-1 у человека вызывает агенезию поджелудочной железы. Функция pdx-1 — «обучающая», на что указывает способность к трансформации печени в поджелудочную железу при гепатической экспрессии гомолога Xenopus pdx-1, спаренного с трансактиватором VP16. Кроме этого, pdx-1 позднее участвует в поддержании идентичности b-клеток и транскрипции гена инсулина. Говоря о роли окружающей мезенхимы, следует упомянуть, что протеин Isl1 LIM-гомеодомена экспрессируется в мезенхиме дорсальной части поджелудочной железы, он необходим для ее развития. Если Isl1 отсутствует, дорсальная часть поджелудочной железы не формируется; при этом вентральная ее часть остается интактной и продолжает развиваться.
Дифференцировка клеток поджелудочной железы: разделение для выполнения экзокринной и эндокринной функцийПосле образования клеток поджелудочной железы и первичного почкования ее зачатка клетки-предшественники энтодермы дифференцируются по двум путям: экзокринному и эндокринному. В последние годы были расшифрованы молекулярные эффекторы, влияющие на эти события и раскрыт механизм распределения клеток между различными органами. Используя в качестве примера опубликованную ранее работу по изучению аллокации нейрональных клеток, ученые исследовали сигнальную систему Notch мутантных мышей и обнаружили, что сигнал Notch является определяющим для последующей дифференцировки клеток в экзокринные или эндокринные. В частности, ранее было известно, что для дифференцировки клеток по эндокринному пути необходим основной «спиральпетля-спираль» (basic helix-loop-helix, bHLH) фактор транскрипции ngn3. ngn3 активирует экспрессию Notch-лигандов Delta, Serrate и Jagged, которые затем активируют сигнальную систему Notch на соседних клетках. Данный сигнал приводит к экспрессии Hes-1, который, в свою очередь, репрессирует ngn и другие гены-мишени. Таким образом, недифференцированная клетка поджелудочной железы, которой уготована роль островковой клетки, будет давать ближайшим клеткам сигнал к выполнению ими экзокринной функции. С помощью указанного механизма, который называют латеральным ингибированием, сигнальная система Notch способствует дифференцировке клеток поджелудочной железы на эндокринные и экзокринные. Недавно проведенные исследования продемонстрировали роль ретиноевой кислоты в дифференцировке клеток. Возможно, ретиноевая кислота действует в направлении, противоположном действию сигнальной системы Notch. Интересно, что результат воздействия ретиноевой кислоты, используемой в качестве лечебного препарата, на дорсальную часть и вентральную часть поджелудочной железы различается. Например, отмечается дифференцировка дорсальной части поджелудочной железы по эндокринному пути, в то время как вентральная часть развивается по экзокринному пути. Эти данные рассматривают как еще одно доказательство существования различных программ развития дорсальной и вентральной частей зачатка поджелудочной железы. – Также рекомендуем “Регуляция дифференциации эндокринных клеток поджелудочной железы” Оглавление темы “Эмбриогенез поджелудочной железы”:
|

Источник